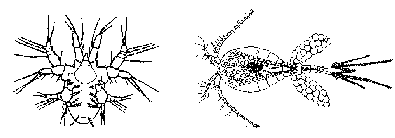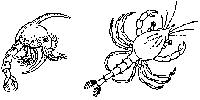 |
My fair larvae |
|
My fair larvae |
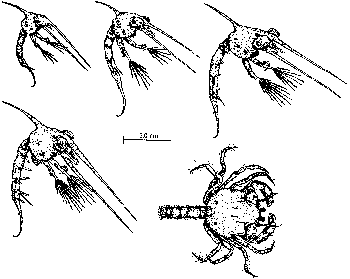
BackConfusing the pathsMany creatures are not sure exactly which of the two paths to follow. They seem to get a bit confused and take them both, combining what appears to be a gradual transformation of tissues and organs at one stage with a more drastic and relatively speedy metamorphosis at another time. The CrabAlmost all saltwater crabs start out life as tiny larvae floating around in the vast ocean. The "typical" female crab usually lays her eggs in winter, often gluing them onto herself. The eggs hatch the following spring into bizarre little larvae called zoeas or crab caterpillars. They paddle about in the surface waters of the sea in a perpetual somersaulting motion, during which they molt three to eight times, depending on the species, producing a number of zoeal stages. Even though all stages have a zoea-like-look about them, they do actually undergo change from one stage to the next, and so the molting process is often called a metamorphic molt (Cameron 1985, p. 103). Somewhere between about three and six weeks, another molt occurs; the zoea loses its spine and produces a radically different larval stage called the megalopa. The megalopa has a squarish head with enormous eyes, two large claws and four pairs of walking legs. It is beginning to look more like a crab. The megalopa still uses its long tail to somersault around for another few weeks before sinking to the bottom. During this period some species molt several times, becoming a little more crablike with each molt. Finally, the young crab tucks in what is left of its tail and becomes an adult, albeit a small one.
Two stages of gradual transformation occur, interrupted by a metamorphosis. The first stage of development consists of the four zoeal phases, while the second stage of development consists of the megalopa phases. Metamorphosis is considered to occur when the zoea turns into a megalopa. The second path: real-life, frog-to-prince storiesWhen it comes to life cycles involving alternating larval and adult phases, practically any variation on the theme you can imagine occurs. Many lifetimes could be spent familiarizing oneself with all the subtle variants superimposed on the macro-variations. Some marine invertebrates that alternate between larval phase and adult phase spend both stages in open water — they never settle down on the bottom. The majority of transformers spend their larval phase in open waters as members of the plankton and their adult lives either crawling across, or attached to, the bottom or, alternatively, as parasites on or inside other animals. Further illustrating the infinite ingenuity of the Creator, some that live as parasites as adults have a bet both ways when it comes to their larvae, living part of their larval life in the plankton and another part as parasites. Enriching the variations even further, others live independently as adults but spend one or more larval stages as parasites! Got all that? Copepod crustaceans of the genus Monstrilla illustrate the last-mentioned pattern. Adults are free-swimmers, as is the first member of its larval stages, known as a nauplius larva. The nauplius tracks down and enters a polychaete worm. Having gained entry by some marvelous means, it breaks into a blood vessel and there grows two or four "arms" that are designed to absorb nutrients from the blood. This phase alone takes in nourishment; it has to build a store of food to last it through all other stages of its life cycle. Neither the adult nor free-swimming nauplius larva eats anything.
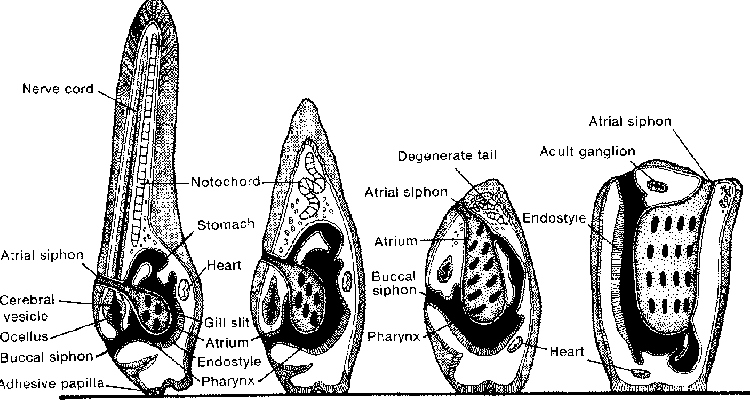
In a small number of cases, the adults live in the open water while the larvae are attached to the sea floor! Small jelly-fish-like creatures known as Hydrozoans illustrate this perverse way of life. The sexually-reproducing adults float or swim in the water column, producing tiny planula larvae that quickly settle and produce colonies of coral-like polyps. Renowned invertebrate expert, Libbie Hyman calls the colonial polyps "a persistent larval stage" (p. 367). Russell-Hunter likewise says, "the relatively massive and extensive hydroid colonies. are by most definitions the larval stages" (p. 577). Just as the change from slimy frog to handsome prince strains credulity so too do the dramatic transformations that occur among many marine invertebrates, such as that of the "tadpole larva" of certain sea squirts, the adults of which are exemplified by those potato-like lumps on the underside of intertidal rocks that "pee" when you squeeze them. Try to imagine the tadpole-like critter in the first picture swimming around happily before attaching itself by its mouth to a rock. Obviously, the changes occurring during metamorphosis in this case are major.
Numerous different kinds of larvae occur, each undergoing its own unique transformation, gradual or rapid, at some stage in life. Space prohibits even a brief survey of all the different kinds and the changes they undergo. The interested reader is advised to find a good zoology book at any library. I can't resist mentioning a couple of my favorites. When found under a rock, ribbon worms, also known as proboscis worms, look like little balls of tangled thread. Put them in an aquarium and, depending on the species, you might see them stretch out into an amazingly long (60 centimeters and more) and thin (a couple of millimeters) worm, often beautifully colored. The pilidium larva of those that have a larval stage undergoes an ingrowth of four or five pockets which eventually fuse around the gut forming a continuous cavity around an inner kernel, separating it from a surrounding husk. The inner mass undergoes a rather complicated metamorphosis into a tiny dense worm which lives like a parasite inside its larval covering. At length the worm is released and drops like a bomb to the sea floor, while the ciliated husk swims off and dies.
The veliger larva of marine snails known as gastropods has a peculiar "skirt" (velum) consisting of two semi-circular lobes studded with long cilia. These cilia do double duty as both propellers and food grabbers. The beating of these long cilia create a current that sucks tinier planktonic motes into a groove lined with stubbier cilia. Within this groove, food particles become entangled in mucus and are spirited to the mouth by synchronized waves of the cilia. During its unique metamorphosis, the entire gut and the incipient shell twist 180 degrees, bringing the anus to just above the head. This process, known as torsion, can occur in as little as three minutes or take days. In some species, torsion "takes place in two 90-degree phases, in which the first is relatively rapid and second more gradual" (Barnes, p. 365).
The intricate design of many larvae renders them worthy of study in their own right. Researchers have found that sea urchin embryos develop "an elaborate crystalline skeleton" (Inoué & Okazaki 1977, p. 83) by a complicated mechanism. The same authors exuberantly declare that, "The simple elegance of the triradiate spicules and the sculptured beauty of the skeletal spicules were wonderful to behold" (p. 86). Oh yes, of course, all hail the power of natural selection. Larval lifeThough one often reads of larvae as being "free swimming", with the ability to swim up or down, one should not interpret that to mean that they can defy currents. Creatures of the plankton are suspended in the water column and are therefore at the mercy of currents, often being carried far from suitable final homes. In spite of the odds, a simple truth of the real world is that those creatures that undergo a larval existence survive quite well, thank you. Density of planktonic larvae varies with latitude and with time of year. Density in tropical seas varies little throughout the year, whereas in temperate zones larvae generally appear when they can take best advantage of the annual Spring Diatom Increase (SDI) which occurs when light and nutrient levels peak, with a resultant dramatic population explosion of photosynthetic algae in spring, at which time algal density increases by 500 to 2000 times winter levels (Russell-Hunter, p. 568), then drops off again as predation outstrips new production. (Since this article was written, scientists have discovered that the Spring Diatom Increase has been largely misunderstood. See Old Theory of Phytoplankton Growth Overturned .) The length of time spent as a free-swimming larva varies from seconds to months. Some sea squirt larvae live in the plankton for seconds (Carefoot, p. 57); hydrozoan planula larvae swim around for a matter of hours at the least or days at the most before attaching to an object and developing into a hydroid colony (Barnes, p. 104); brittle star larvae are thought to spend several weeks in the plankton before settling as juveniles, while "the larvae of some species [of coral] are capable of remaining for months in the surface waters" (Endean 1982, p. 61). Many starfish larvae drift for two months before settlement. By contrast with those larvae that spend considerable time as members of the plankton and swim upwards towards light, rapid settlers are not attracted to light.
Not surprising to those familiar with the creation, exceptions occur to every general statement one can make. Rather than entering the plankton, larvae of sea spiders (Pycnogonids) attach themselves to their father's legs or, in other cases, live as parasites on hydroids or corals! (Barnes, p. 508). Whatever you can dream up, it happens — somewhere. Modern researchers continue to discover amazing facts, such as that some species produce two different kinds of larvae — a non-feeding kind that settles very quickly by swimming downwards immediately upon hatching and a feeding kind that lives in the plankton for a period of time before settling. Why? Who can say? Researchers suggest that such an arrangement enhances "dispersal and settlement into patchy adult habitats" (Krug and Zimmer). Whatever that might mean. A universal truth of larval life is that mortality rates are extremely high, particularly during early larval life and later upon settling. To compensate for the staggering losses, huge numbers are produced; some starfish release up to 200 million eggs in one laying episode. The Creator of all figured out the specific vicissitudes that particular kinds of larvae would undergo during all phases of their existence, calculated the numbers precisely, and got it right every time when it came to determining the balance between the number that must be produced to keep the species going but not for it to rule the world. FlotationThe richest pastures of the seas and oceans are found in the photic zone, the top 100 meters of the sea, and particularly the very uppermost few meters where single-celled algae such as diatoms, which form the basis of a rich food chain, are found. Larvae must stay near the surface or starve. The mechanics of flotation of larvae has presented researchers with some headaches. In 1954 Coker declared that they were "not a fully-solved problem" (p. 108). Why don't larvae sink? Experts generally attribute their buoyancy, enabling them to stay in the food-rich upper layers, to factors such as,
Undoubtedly other hitherto unknown factors will come to light with time. Oh the genius of the Creator. Returning homeHow do larvae that have been subject to the whims of currents and washed many miles from home return to their place of "birth"? Blue crabs of Chesapeake Bay leave the shallow waters of estuaries and muddy shores for the ocean to release their eggs (Cameron, p. 103). Obviously, tiny larvae produced by inshore creatures may be swept way out to sea or drift long distances from home. How do blue crab larvae get back to the Bay, and shoreline larvae back to the shore? Well, it seems as if survival depends upon caprice. Most never make it to a suitable location; flukes of current and turbulence, however, always ensure enough get to where they can make a home. Enough is enough. Many barnacles illustrate convincingly the role played by chance in the survival of their larvae. To survive, mature larvae must find themselves in the wave swash "when the tide is high, and only when it is high" (Carefoot, p. 32). Tough luck for those ready to settle when the tide is out! For some years, oceanographers struggled with the "long-standing conundrum" (Copley 1998, p. 32) of how bizarre marine organisms found around deep-sea vents were able to colonize lifeless distant vents. These vents, found along the chains of undersea volcanoes along subduction zones, spew out hot seawater which contains minerals that grow into chimneys and nourish rich communities of bizarre invertebrates. Vents come and go; old ones are wrecked by earthquakes, or they become clogged with mineral deposits, spelling doom to their inhabitants. For deep-sea vents to be populated, some mechanism must ensure that new vents are populated, even when they are several hundred kilometers from the nearest live one. God had to solve the problem; how did He do it? In the last years of the twentieth century, researchers discovered the existence of giant swirling whirlpools hundreds of meters above the ocean floor generated by the vents. Copley says that, "scientists have discovered that these spinning masses of water can break free and wheel across the underwater landscape like flying saucers, carrying heat, chemicals and even vent animals heading for new pastures. Many of the animals around the vents don't move much as adults, but they do have tiny offspring that could be carried away by currents. Perhaps spinning eddies could transport these larvae in concentrated patches from one vent site to another" (p. 31-32). One expert reckons that as water spews out of the vents, it creates a spreading plume that would turn into a rotating lens of water roughly 200 meters thick and 2 kilometers across. It picks up larvae, breaks free, spins away like a toy top, and may eventually stumble upon a newly-formed vent, seeding it with life. SettlingIf the design of larvae warrants study, their behavior invites even more investigation. Though getting within spitting distance of a suitable final resting place for larvae of stay-at-home parents is a matter of serendipity, choosing the precise location is done with as much attention to detail as by executives of a fast-food chain looking for a new outlet site. For larvae of bottom-dwellers, instinct has to reverse itself in one critical respect — larvae are attracted to light during early larval life, ensuring they actively stay near the surface where their food is found, then switch to seeking shade as the time for metamorphosis approaches. (Some react, instead, to gravity or pressure). Larvae of those creatures found only in the intertidal zone remain attracted to light to the bitter end, enabling them to colonize the upper reaches of the shore.
For most species, the challenge of finding a suitable home is far more complex than merely "heading down", requiring far more discriminatory capabilities than reacting to light or gravity. I often visit a richly-inhabited intertidal rock platform jutting out from the shore for about 200 meters at Penguin, Tasmania. You don't have to be a Sherlock Holmes to note that many creatures are almost always found in a given location or zone. Some are found at the high tide line only. Feather stars, I have observed, are invariably found on the extreme west side at or below low tide mark; from memory, I have never found one on the eastern side. I suspect that their behavior is governed by factors similar to one that influences site selection in Atlantic barnacles: "… larvae of [Atlantic] barnacles are not stimulated to settle in conditions of slow current" (Carefoot, p. 31). Arriving at a suitable general location by chance, how do they then select the precisely-right place? Many instances of ability to find the right habitat almost beggar belief. The stalked barnacle, Conchoderma, is always found living on another barnacle, Coronula diadema, which is invariably found attached to the underside — not the back — of an Antarctic humpback whale (Carefoot, p. 59). We will conclude this article with some quotes that clearly express the remarkable discriminatory power of larvae: Most, if not all, larvae of bottom-dwelling marine invertebrates have relatively sophisticated discriminatory capabilities, allowing them — for a short time, at least, before adopting the adult mode of life — to crawl over and test the substrate for its suitability as a settlement site. Not until the 1920s did biologists begin to realize that larval settlement was not the hit-or-miss affair that they had imagined, but they took several more decades to appreciate just how complex the factors are that stimulate settlement and metamorphosis, and how sensitive the larva must be to these stimuli (Carefoot, p. 47 & 60). Endean, speaking about coral larvae, says this: Little is known about how the larvae choose a place to settle. The presence of numerous predatory organisms makes settling a hazardous process. In some cases the presence of coralline algae appears to be a prerequisite for successful settling of coral larvae. Many appear to settle near their parents. (1982, p. 61). Barnes on gastropod larvae says this: Settling sites are of critical importance in the survival of the larvae, and many species can delay metamorphosis until specific types of substrates can be reached. Some nudibranchs, for example, must contact certain species of hydroids, bryozoans, or ascidians before metamorphosis will occur. Metamorphosis appears to be induced by chemical rather than physical characteristics of the substratum (p. 365). In short, a staggering array of cues and factors are used by larvae of different species to find those spots conducive to — nay, essential to — the good life. What is more, larvae somehow know to space themselves out according to their adult size! Oh the wisdom of the supreme Intelligent Designer. Who could possibly believe that such sophistication came about because a tiny speck of infinite mass exploded?
|
References and notes Allen, Larry G. May 1983, Larval Development of the Northern Clingfish, Gobiesox maeandricus, Copeia Ballinsky, B. I. 1970, An Introduction to Embryology, W. B. Saunders Company, Philadelphia Barnes, Robert D. 1974, Invertebrate Zoology, W. B. Saunders Company, Philadelphia Cameron, James N. May 1985, Molting in the Blue Crab, Scientific American Carefoot, Thomas 1977, Pacific Seashores, University of Washington Press, Seattle Coker, R. E. 1954, This Great and Wide Sea, Harper Torchbooks, New York Copley, Jon 12 December 1998, Going for a Spin, New Scientist Endean, Robert 1982, Australia's Great Barrier Reef, University of Queensland Press Gilbert, S. F. and Raunio, A. M. 1997, Embryology: Constructing the Organism, Sinaeuer Associates, Inc. Publishers, Sunderland Gould, S. J. December 1986, Glow, Big Glowworm, Natural History Hyman, L. H. The Invertebrates: Coelenterata, McGraw-Hill, New York Inoué, S., and Okazaki, K. April 1977, Biocrystals, Scientific American Krug and Zimmer, www.biolbull.org/cgi/content/abstract/207/3/233 Wigglesworth, V. B. 1959, Metamorphosis and Differentiation, Scientific American Offprint 63 Williamson, D. I. 1992, Larvae and Evolution: Toward a New Zoology, Chapman and Hall, New York Young, J. Z. 1962, The Life of Vertebrates, Oxford University Press, New York & Oxford |
Further reading |
Dawn to Dusk publications
|
Other printed material |
On the web |
Any zoology text will have plenty of interesting information. Just check "larvae" and "metamorphosis" in the index. One highly recommended book is, Embryology: Constructing the Organism, by Gilbert and Raunio. An interesting short article is S. J. Gould's, Glow, Big Glowworm, Natural History magazine, December 1986 |
http://www.microscopy-uk.org.uk/mag/artjul00/echino.html — has some fine pics of larvae An excellent Natural History magazine article available online: |
| Edited and expanded copies of this article, in reprint pamphlet form, can be purchased by going to the reprints order page. As well as reprints, Dawn to Dusk offers books in printed form and on CD-ROM. We mail to anywhere in the world! For more information on what is available, prices, and how to order, click the icon. |